ДНК
Дезоксирибонуклеї́нова кислота́ (ДНК) — один із двох типів природних нуклеїнових кислот, що забезпечує зберігання, передачу з покоління в покоління і реалізацію генетичної програми розвитку й функціонування живих організмів. Основна роль ДНК в клітинах — довготривале зберігання інформації про структуру РНК і білків.
Загальний опис
Загальний опис
У клітинах еукаріотів (наприклад, тварин, рослин або грибів) ДНК міститься в ядрі клітини в складі хромосом, а також в деяких клітинних органелах (мітохондріях і пластидах). У клітинах прокаріотів (бактерій і архей) кільцева або лінійна молекула ДНК, так званий нуклеодії, міститься в цитоплазмі і прикріплена зсередини до клітинної мембрани. У них і у нижчих еукаріотів (наприклад дріждів) зустрічаються також невеликі автономні кільцеві молекули ДНК, так звані плазміди. Крім того, одно- або дволанцюгові молекули ДНК можуть утворювати геном ДНК-вірусів.
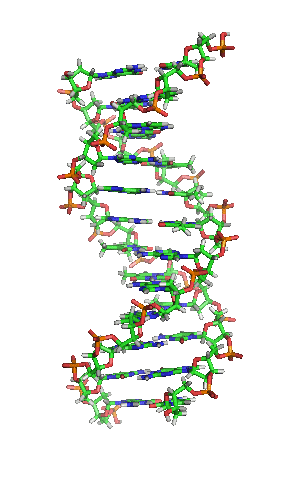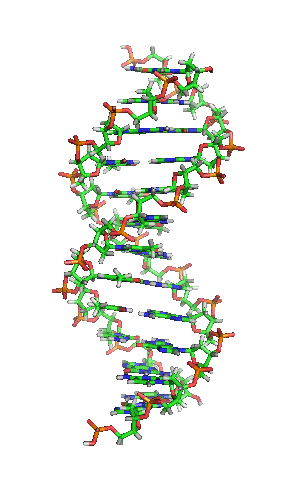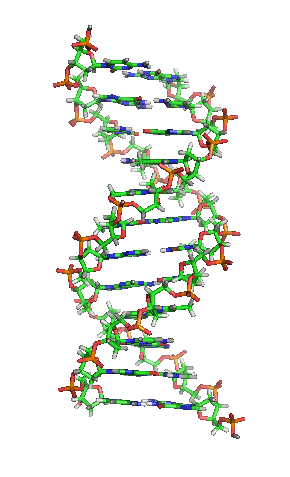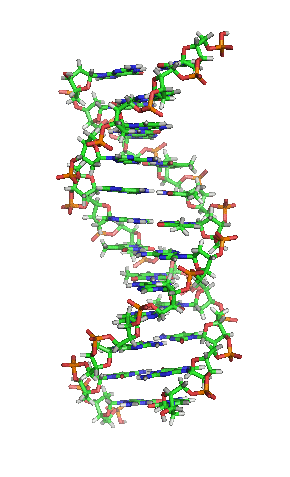
З хімічної точки зору ДНК — це довга полімерна молекула, що складається з послідовності блоків —нуклеотидів. Кожний нуклеотид складається з азотистої основи, цукру (дезоксирибози) і фосфатної групи. Зв'язки між нуклеотидами в ланцюгу утворюються за рахунок дезоксирибози і фосфатної групи. У переважній більшості випадків (окрім деяких вірусів, що містять одноланцюгові ДНК) макромолекула ДНК складається з двох ланцюгів, орієнтованих азотистими основами один проти одного. Ця дволанцюгова молекула утворює спіраль. У цілому структура молекули ДНК отримала назву «подвійної спіралі».
У ДНК зустрічається чотири види азотистих основ (аденін, гуанін, тимін і цитозин). Азотисті основи одного з ланцюгів сполучені з азотистими основами іншого ланцюга водневими зв'язками згідно з принципом комплементарності: аденін з'єднується тільки з тиміном, гуанін — тільки з цитозином.
Послідовність нуклеотидів дозволяє «кодувати» інформацію про різні типи РНК, найважливішими з яких є матричні (мРНК), рибосомні (рРНК) і транспортні (тРНК) та інші некодуючі РНК. Всі ці типи РНК синтезуються у процесі транскрипції на матриці ДНК, тобто за рахунок копіювання послідовності ДНК у послідовність макромолекули РНК, за допомогою принципу комплементарності. Деякі види РНК, такі як мРНК, тРНК, рРНК за допомогою малих ядерних РНК беруть участь у Біосинтезі білків(процесах транскрипції, сплайсингу і трансляції). Крім кодуючих послідовностей, ДНК клітини містить некодуючі послідовності, що виконують регуляторні і структурні функції, або не виконують ніяких функцій. Ділянки кодуючих послідовностей разом із регуляторними ділянками називаються генами. Сукупність всіх генів, регуляторних послідовностей, некодуючих послідовностей, тобто вся нуклеотидна послідовність ДНК незалежно від її функцій, формує геном організму.
У геномах еукаріотів містяться також довгі послідовності без очевидної функції (некодуючі послідовності). Також у складі геному досить поширені генетичні паразити — транспозони і вірусні або схожі на них послідовності. Проте організм може використовувати транспозони для виконання певних функцій, також транспозони можуть впливати на еволюцію генів.
Розшифровка структури ДНК, виконана в 1953 році, стала одним з поворотних моментів в історії біології. За видатний внесок у це відкриття Френсісу Кріку, Джеймсу Ватсону і Морісу Вілкінсу була присуджена Нобелівська премія з фізіології або медицини 1962 року.
Структура молекули
Нуклеотиди
Дезоксирибонуклеїнова кислота є біополімери, мономерами якого є нуклеотиди. Кожен нуклеотид складається із залишку фосфорної кислоти, приєднаного за 5'-положенням до цукрудезоксирибози, до якого також через глікозидний зв'язок(C—N) за 1'-положенням приєднана одна з чотирьох азотистих основ. Саме наявність характерного цукру і складає одну з головних відмінностей між ДНК і РНК, зафіксовану в назвах цих нуклеїнових кислот (до складу РНК входить цукор рибоза). На малюнку показано чотири основи та один із нуклеотидів — дезоксаденозинмонофосфат, утворений приєднанням аденіну до дезоксирибози й фосфату.
Дезоксирибоза, що входить в склад молекули ДНК, знаходиться переважно в С2'-ендо конформації, на відміну від С3'-ендо конформації рибози в молекулах РНК: відсутність OH-групи у 2 положенні дезоксирибози робить С2'-ендо конформацію можливою. Це надає ДНК структурної міцності і ригідності, на відміну від молекули РНК.
За структурою молекул азотисті основи, що входять до складу нуклеотидів, розділяють на дві групи: пуринові(аденін [A] і гуанін [G]), утворені сполученими п'яти- і шестичленним гетероциклами і піримідинові (цитозин [C] і тимін [T]) — утворені одним шестичленним гетероциклом.
Як виняток, наприклад, у бактеріофага PBS1, в ДНК зустрічається п'ятий тип основ — урацил (U), піримідинова основа, що зазвичай входить до складу РНК замість тиміну і відрізняється від тиміну відсутністю метильноїгрупи на кільці. Слід зазначити, що тимін і урацил не так строго приурочені до ДНК і РНК відповідно, як це вважалося раніше. Так, після синтезу деяких молекул РНК значне число урацилів у цих молекулах метилюєтся за допомогою спеціальних ферментів, перетворюючись на тимін. Це відбувається в транспортних і рибосомних РНК.
Подвійна спіраль
Полімер ДНК має досить складну структуру. Нуклеотиди ковалентно сполучені між собою в довгі полінуклеотидні ланцюги. Ці ланцюги в переважній більшості випадків (окрім деяких вірусів, що мають одноланцюговий ДНК-геном), у свою чергу, попарно об'єднуються за допомогою водневих зв'язків у структуру, що отримала назву подвійної спіралі. Фосфатні групи формують фосфодіестерні зв'язки між третім і п'ятим атомами вуглецю сусідніх молекул дезоксирибози, в результаті взаємодії між 3-гідроксильною групою (3-ОН) однієї молекули дезоксирибози і 5-фосфатною групою (5-РО3) іншої. Асиметричні кінці ланцюга ДНК називаються 3' (три-прайм) і 5' (п'ять-прайм). Полярність ланцюга грає важливу роль при синтезі ДНК (подовження ланцюга можливе тільки шляхом приєднання нових нуклеотидів до вільного 3'-кінцю).
Як вже було сказано вище, у переважної більшості живих організмів ДНК складається не з одного, а з двох полінуклеотидних ланцюгів. Ці два довгі ланцюги закручені один навколо іншого у вигляді подвійної спіралі, що стабілізується водневими зв'язками, які утворюються між повернутими один до одного азотистими основами ланцюгів, що входять до неї. У природі ця спіраль зазвичай правозакручена. Напрями від 3'-кінця до 5'-кінця в двох ланцюгах, з яких складається молекула ДНК, протилежні (ланцюги «антипаралельні» один одному).
Ширина подвійної спіралі в її найпоширенішій В-формі становить 2,2 — 2,4 нм, а довжина кожного нуклеотида 3,3 Å (0,33 нм). Довжина всієї молекули залежить від виду організму, та може складати від десятків мікрон у деяких вірусів до кількох метрів (в одній хромосомі) у деяких рослин. Подібно до того, як в гвинтових сходах збоку можна побачити сходинки, на подвійній спіралі ДНК в проміжках між фосфатним остовом молекули можна бачити ребра основ, кільця яких розташовані в площині, перпендикулярній до подовжньої осі макромолекули.
Подвійна спіраль ДНК має декілька параметрів конформації, які можна характеризувати:
- шифт, слайд, райз — характеризують зсув однієї пари основ відносно іншої без зміни куту площини пар основ, так, що вони залишаються паралельними;
- твіст, ролл, тилт— характеризують повороти площин однієї пари основ відносно іншої.
Жолобки
У подвійній спіралі розрізняють малий (12 Å) і великий (22 Å) жолобки (борозенки). Білки, наприклад, фактори транскрипції, які приєднуються до певних послідовностей в дволанцюговій ДНК, зазвичай взаємодіють з краями основ у великому жолобку, де вони доступніші.
ДНК може мати триланцюгову форму при утворенні зв'язків з третім ланцюгом ДНК (або РНК) у великому жолобку через формування окрім канонічних ватсон-кріківських пар, третю хугстинову водневу взаємодію.
Біологічна роль
ДНК є носієм генетичної інформації, записаної у вигляді нуклеотидної послідовності за допомогою генетичного коду. З молекулами ДНК зв'язані дві основні властивості живих організмів — спадковість і мінливість. У ході процесу, що називається реплікацією ДНК, з початкового, материнського, ланцюга, утворюються дві копії ДНК, які успадковуються дочірніми клітинами при поділі. Клітини, що утворилися таким чином, будуть генетично ідентичними. Потрібна для клітинної життєдіяльності генетична інформація зчитується при експресії генів. У багатьох випадках вона використовується для біосинтезу білків у процесах транскрипції (синтезу молекул РНК на матриці ДНК) і трансляції (синтезу білків на матриці РНК).
Послідовність нуклеотидів «кодує» інформацію про різні типи РНК: кодуючі — матричні (мРНК) — та некодуючі — рибосомні (рРНК), транспортні (тРНК), каталітичні та інші. Всі ці типи РНК синтезуються на основі ДНК у процесі транскрипції. Їхня роль у біосинтезі білків та інших процесах життєдіяльності клітини різна. Матрична РНК містить інформацію про послідовність амінокислот у білку, рибосомальні РНК служать основою для рибосом (складних нуклеопротеїнових комплексів, основна функція яких — збірка білка з окремих амінокислот на основі мРНК), транспортні РНК доставляють амінокислоти до місця збірки білків — в активний центр рибосоми, що рухається по мРНК, проте синтезуються також некодуючі РНК, які можуть виконувати різноманітні регуляторні функції.
Реплікація
Поділ клітини необхідний для розмноження одноклітинних і росту багатоклітиннних організмів, але до поділу клітина повинна подвоїти геном, щоб дочірні клітини містили ту ж генетичну інформацію, що і початкова клітина. ДНК подвоюється у процесі реплікації, що протікає за напівконсервативним механізмом: два ланцюги розділяються, і потім кожна комплементарна послідовність ДНК відтворює для себе пару за допомогою ферменту ДНК-полімераза. Цей фермент будує полінуклеотидний ланцюжок, знаходячи правильний нуклеотид через комплементарне спаровування основ і приєднуючи його до зростаючого ланцюга. ДНК-полімераза, що здійснює більшу частину синтезу не може розпочати синтез нового ланцюга, а тільки нарощує вже існуючий, тому вона потребує наявності праймерів, ділянок ДНК, синтезованих за допомогою спеціальної РНК-полімерази праймази. Оскільки ДНК-полімерази можуть будувати ланцюжок тільки у напрямку 5' → 3', для копіювання антипаралельних ланцюгів використовуються складні механізми з залученням великої кількості білків, і цей ланцюг копіюється переривчасто, невеликими ділянками — фрагментами Окадзакі, довжиною близько 200 bp.
Історія дослідження ДНК
Історія дослідження ДНК
ДНК була відкрита Іоганном Фрідріхом Мішером у 1869 році. Спочатку нова речовина отримала назву «нуклеїн», а пізніше, коли Мішер виявив у неї кислотні властивості, її назвали нуклеїновою кислотою. Біологічна функція нововідкритої речовини була неясна, і довгий час ДНК вважалася запасником фосфору в організмі. Більш того, навіть на початку 20 століття багато біологів вважали, що ДНК не має стосунку до передачі інформації, оскільки будова молекули, на їхню думку, була дуже одноманітною і не могла містити закодовану інформацію.
Поступово було доведено, що саме ДНК, а не білки, як вважалося раніше, є носієм генетичної інформації. Одними з перших вирішальних доказів стали експерименти О. Евері, Коліна Мак-Леода і Маклін Мак-Карті (1944 рік) з трансформації бактерій. Їм вдалося показати, що за так звану трансформацію (придбання хвороботворних властивостей нешкідливою культурою у результаті додавання до неї мертвих хвороботворних бактерій) відповідає виділена з пневмококів ДНК. Експеримент американських учених Алфреда Хершу і Марти Чейз (1952 рік) з міченими радіоактивними ізотопами білками і ДНК бактеріофагівв показали, що в заражену клітину передається тільки нуклеїнова кислота фага, а нове покоління фага містить такі ж білки і нуклеїнову кислоту, як і початковий фаг.
До 50-х років 20 століття точна будова ДНК, як і спосіб передачі спадкової інформації, залишалася невідомою. Хоч і було напевно відомо, що ДНК складається з кількох ланцюгів, що у свою чергу складаються з нуклеотидів, ніхто не знав точно, скільки цих ланцюгів і як вони сполучені.
Структура подвійної спіралі ДНК була запропонована Френсісом Кріком і Джеймсом Ватсоном у 1953 році на основі рентгеноструктурних даних, отриманих Морісом Вілкінсом і Розаліндою Франклін, і "правил Чаргаффа", згідно з якими в кожній молекулі ДНК дотримуються строгі співвідношення, що зв'язують між собою кількість азотистих основ різних типів. Пізніше запропонована Ватсоном і Кріком модель будови ДНК була доведена, а їхня робота відмічена Нобелівською премією з фізіології і медицини 1962 року. Серед одержувачів не було Розалінди Франклін, що померла на той час, оскільки премія не присуджується посмертно.
У відомій доповіді 1957 року Крік окреслив основи так званої "Центральної догми" молекулярної біології, яка передбачає взаємовідношення між ДНК, РНК і білками, та сформулював "адаптерну гіпотезу". Остаточне підтвердження механізму копіювання, запропонованого на основі спіральної структури, було отримане в 1958 році за допомогою експерименту Мезельсона-Сталя. Подальші роботи Кріка і його лабораторії показали, що генетичний код засновується на трійках основ, що не перекриваються, кодонах, що пізніше дозволило Гару Гобінду Хорані, Роберту Голлі і Маршаллу Ніренбергу розшифрувати генетичний код. Ці відкриття позначають початок молекулярної біології.


Комментарии
Отправить комментарий